
1.1细菌的群体感应现象
20世纪60年代以来,随着Nealson K H等在海洋细菌费氏弧菌(Photobacterittm fischeri,曾用名Vibro fiseheri)和夏威夷弧菌(Vibro haveryi)中发现了由群体感应控制的生物发光现象及其深入研究,人们逐渐的认识到细菌之间具备信息交流能力。细菌间的交流并不像人类间的交流通过“声波”,而是依靠某一种或多种信号分子相互交谈,细菌之间可以根据这些信号分子的浓度的高低来感受周围环境中自身或其他细菌的数量变化,并通过信号分子发出信号, 启动菌体中相关基因的表达, 改变和协调它们之间的行为, 共同展示出它们的某些生理特性, 从而表现出单个细菌无法从事的某些生理功能和调节机制。这一调控系统被称为细菌的群体感应调节(quorum sensing,QS)。
不同的细菌在繁殖过程中分泌不同的信号分子来调控基因的表达,这种信号分子一般可以自由的从胞内扩散到胞外。当这种信号分子达到一定的阈值时,细菌可以感知到菌群的细胞密度,启动某些基因的表达。据已有的研究表明,革兰氏阴性菌通过产生高丝氨酸内酯类 (acyl—homoserine lactonase,AHLs)作为细胞间的信号分子,该信号分子以细胞密度依赖的方式调控生理特性的表达;革兰氏阳性菌利用一些小肽作为QS的信号分子来感知种内自身种群数量,协调多种基因的表达;还有一类信号分子是一种呋喃酰硼酸二酯,即自体诱导子,它在革兰氏阴性菌和阳性菌中均存在,被用来感知种间细菌数量来调控细菌自身行为[3]。
对于革兰氏阴性菌的AHLs类信号分子的特点为:均含有一个高丝氨酸内酯环和4~14个碳不等的酰基侧链,该特征性内酯环结构在质谱鉴定过程中不会破坏。在本实验中,实验菌种E.coli Top10F'菌株产生的信号分子为相对分子量为213,分子式为C10H15NO4,化学式为3-oxo-C6-HSL(若无特殊说明,一下提及的信号分子均为该分子),分子结构如图1-1所示:
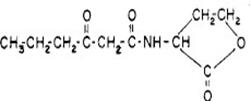
图1-1 C10H15NO4分子结构图
Fig 1-1 Molecular structure of C10H15NO4
1.2革兰氏阴性细菌的群体感应系统
AHL分子都含有一个高丝氨酸内酯环,不同的AHL分子含有不同的酰化支链。酰化支链的差异是AHL群体感应系统保证种内特异性的分子基础。AHL分子的作用机制是相似的:AHL分子一般是在Lux I蛋白酶的催化下,以硫腺苷甲硫氨酸(S-adenosylmethionine,SAM)和酯酰基载体蛋白(acyl-axyI carrier protein,ACP)为底物合成的。当细菌密度增大(达到107个细菌/ml时),AHL分子在环境中累计到一定浓度,AHL分子进入细胞与相应的受体Lux R结合。Lux R可以与靶基因的调控元件结合,是一种转录激活物。Lux R-AHL复合物一般以二聚体的形式结合于靶基因启动子区域从而启动基因的表达。AHL群体感应系统感知种内数量的特异性主要由以下两个方面决定:一方面是Lux I蛋白酶催化底物的专一性,它只能识别含有特殊酰化侧链的AHL;另一方面是受体Lux R与AHL分子结合的特异性。AHL分子能调控细菌许多致病基因的表达。AHL感应机制示意如图1-2所示:
图1-2 AHL感应机制
Fig 1-2 Sensing mechanism of AHL
1.3本课题的研究思路
合成生物学是生物科学在二十一世纪刚刚出现的一个分支学科,与基因工程把一个物种的基因延续、改变并转移至另一物种的作法不同,合成生物学的目的在于建立人工生物系统(artificial biosystem)。它是在阐明并模拟生物合成的基本规律的基础上,人工设计并构建新的、具有特定生理功能的生物系统,从而建立药物、功能材料或能源替代品等的生物制造途径。例如把网络同简单的细胞相结合,可提高生物传感性,帮助检查人员确定地雷或生物武器的位置。再如向网络加入人体细胞,可以制成用于器官移植的完整器官。 一个基于群体感应的合作质粒的构建(2):http://www.youerw.com/shengwu/lunwen_18098.html