
G蛋白将细胞外的第一信使激素和细胞内的第二信使cAMP联系起来。G蛋白介导的信号转导途径较为保守。G蛋白位于质膜内侧,当细胞转导胞外信号时,首先胞外基团与质膜上的G蛋白偶联受体(GPCRs)结合,受体活化后激活质膜内侧的G蛋白,G蛋白再激活其下游的各种效应器,在细胞内生成第二信使,从而将信号传递下去,调节生物的生长发育。
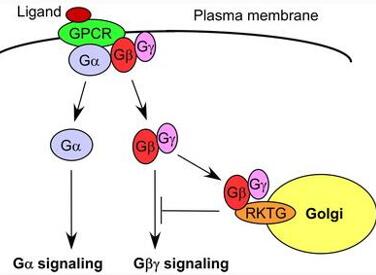
图1:G蛋白偶联受体信号转导调控机制(鞠冠华等,2012)
G蛋白在很多方面调节着植物的生长发育。在植物中,G蛋白参与信号转导、几种植物激素调节以及抗病性和抗逆性相关基因表达等。异源G蛋白调节能够调节植物地上部分,根部及表皮的生长发育。拟南芥gpa1突变体表现为植株矮小,花和果荚变小,结实量变低的表型[2]。虽然其主根没有变化,但侧根变少;与gpa1相比,拟南芥Gβ亚基突变体agb1表现出更严重的发育缺陷,其植株更加矮小,花和果荚更小更短。水稻d1突变体、玉米ct2突变体表现为更严重的发育缺陷,其植株高度仅为野生型的25 %-50 %,根长略有变短。水稻Gβ至今没有功能缺失突变体,推测可能Gβ突变导致胚胎致死,其RNAi植株表现出叶鞘和叶片变窄的表型,而过表达则表现为分蘖多,叶鞘少的现象。在大豆中,Gα蛋白能够直接与节瘤因子NFR1互作正调控根瘤的形成。NFR1受体能够磷酸化并激活RGS,激活的RGS能够使Gα蛋白文持在失活状态,从而使根瘤能够正常发育。拟南芥功能缺失突变体agb1能够恢复bri1造成的植株矮小表型。拟南芥AGB1能够与RLK蛋白家族的ZAR1蛋白互作调节受精卵的不对称分裂。AGB1与ERECTA关联调节拟南芥果荚发育。玉米FEA2能够与CT2(拟南芥GPA1同源蛋白)互作调节玉米SAM干细胞增殖。G蛋白还能影响植物气孔发育。拟南芥gpa1突变体中气孔密度比野生型降低20–30 %,过表达之后,下胚轴表皮中的气孔密度变为原来的5倍,agb1突变体中叶片气孔密度提高25 %,并表现出气孔聚集现象,gpa1和agb1对气孔发育的影响表现为加性效应[3]。
在植物中,G蛋白参与各种信号转导系统,包括几种植物激素调节、抗病性和抗逆性相关基因表达。G蛋白信号转导是一个复杂的调控网络,RLKs和RGS是G蛋白信号转导过程的重要调节因子[4]。植物拥有上百个膜定位的RLKs,RLKs具有非常保守的结构,N端胞外结构域用于感知信号,1-3个跨膜结构域使之结合与细胞膜上,胞内激酶结构域用于磷酸化下游蛋白。与动物中RLKs拥有的酪氨酸激酶活性不同,植物中的RLKs具有丝/苏氨酸激酶活性。拟南芥AtWNK8 (一种RLK)能够磷酸化AtRGS1,促进其内吞,因此是G蛋白信号转导的重要调控因子。除了这些互作证据外,目前并没有更深入研究去探索RLKs调控G蛋白循环的具体机制。RGS是另一类G蛋白信号调节因子。在质膜上,RGS降低了非活性GPA1被激活的速率,当RGS被内吞后,GPA1的激活速率被提高。除了RLKs,RGS外,位于G蛋白信号下游的MAPKs在该信号转导途径中也起着十分重要的作用。串联MAPKs由三种类型,包括MAPKKKs,MAPKKs,MAPKs,他们通过依次磷酸化激活其活性,从而调控G蛋白信号。近来的研究表明,拟南芥RACK1A、RACK1B、RACK1C编码的RACK1蛋白位于G蛋白信号上游,是第一被发现的MAPKs支架蛋白,RACK1的发现,也从侧面印证了MAPKs对G蛋白信号的调控作用[5]。
G蛋白在植物激素调节中也起着重大作用。GPA1 和GCR1在ABA调节的逆境胁迫调节中起着重要的作用。通过对gpa1,gcr1,gpa1 gcr1突变体的表型比较发现,G蛋白在某些程度上调节了BR和GA调节的种子萌发,即G蛋白信号与BR和GA信号之间存在Cross-talk。通过遗传学和分子生物学手段证明,油菜素內酯能够诱导乙烯的生物合成,进而激活Gα蛋白活性,促进H2O2和NO积累,最终导致气孔关闭。拟南芥Gα功能缺失突变体gpa1对ABA诱导的钾离子通道抑制和慢型阴离子通道活性不敏感,导致ABA诱导的气孔关闭被抑制。Gβ亚基突变体 agb1与此相同,表明Gα、Gβ亚基对ABA诱导的气孔开度有重要影响。此外,近来的研究表明,AGG3与GPA1/AGB1共同作用调节ABA影响的的气孔开度[7]。PP2C是ABA信号转导通路中的必要成分,研究表明,拟南芥AGB1能够与PP2C52在细胞膜上互作[8]。以上结果表明,G蛋白参与了ABA信号转导途径。综上所述,G蛋白信号转导与植物激素信号转导之间存在复杂的交联。 IDR1维持巴西陆稻(BXLD)耐热能力的表型研究及机制初探(3):http://www.youerw.com/shengwu/lunwen_19391.html