
在对众多的抗性基因筛选过程中,经历比较长的一段时间的工作后,发现Sw-5抗性基因是具有较广谱的抗性,且抗性比较稳定,易于作为基因工程所需的原材料,使植物拥有对番茄斑萎病毒的特异抗性。Sw-5是显性基因,位于番茄第九条染色体的长臂且接近端粒的地方,在对该基因进行重复试验观察分析后,能够发现在这个位点处有两个高度同源的候选基因,被命名为 Sw-5a和 Sw-5b,经过转基因可以总结出结果, Sw-5b 是介导抗性的要害基因,该基因属于 CC (coiled coil)- NB( nucleotide binding)- ARC (adaptor shared among APAF-1, R proteins, and CED-4)-LRR (leucine-rich repeat)的R基因类型抗性基因[3-4],由于抗性基因的存在,引起细胞防御机制的启动,侵染位点会自动产生过敏性细胞死亡反应(hypersensitive response,HR)[5],在植物表面也会表现出相应的症状。
番茄斑萎病毒有着布尼亚病毒科侵染人和其他动物的病毒类似的特点。番茄斑萎病毒是具有三个基因组的多分体RNA病毒,病毒被囊膜包被,三个RNA链可由大到小分类,而且每个RNA链的末端都存在有约65个核苷酸互补序列,序列三级结构折叠后形成形似锅柄状的构造。其中,中等大小的M RNA链可以编码非结构蛋白NSm,同属于布尼亚病毒科的其它三种动物病毒不能编码类似NSm的移动蛋白,因此,NSm被研究者认为长期自然选择过程中而适应植物的结果[6]。
利用免疫标记等方法,Kormelink等(1994)初次提取和纯化了NSm蛋白,验证了了NSm参与了病毒在植物细胞中的胞间运动(cell-to-cell movement)[7] ,Storms等(1995)将NSm载体构建转入黄花烟原生质体和Tn细胞,荧光标记分析原生质体和昆虫细胞表面,发现NSm能形成管状结构[8],Lewandowski和Adkins(2005)使用运动缺陷型的烟草花叶病毒(TMV)作为原材料,重组NSm,发现能够互补运动缺陷的TMV在细胞间运动并可以长距离运输,在原生质体中使用显微观察发现,发现有与上述相似的管状形状的结构存在[9]。这些特征的发现与侵染性豇豆花叶病毒(CPMV)运动蛋白(MP)的功能具有明显的的相似性[10],充分证明了NSm蛋白具有植物病毒运动蛋白的特征,因而被视作番茄斑萎病毒属病毒的运动蛋白。
Sw-5是染色体中的一个单基因显性遗传位点,能够对多种番茄斑萎病毒属的病毒产生抗性,包含TSWV、GRSV、 TCSV和GB-NV。经过研究,发现Sw-5可以与1个RFLP分子标记CT220密切的连锁在一起[11],以CT220标记为探针得到的1个包含Sw-5的长约400 kb大小的YAC克隆TY257,经过染色体步移的措施,在黏粒文库中将Sw-5这个基因定位在含有9个堆叠黏粒克隆的100 kb的染色体片段中。FOLKERTSMA等[12]应用含有Sw-5抗性基因的番茄纯系为基础构建BAC文库,并使用2个RFLP分子标识CT220和SCAR421的序列为模板设计引物,通过克隆后得到一条长度约250 kb的重叠群,以抗性基因的NBS保守序列再次设计合并引物,克隆得到3个独立的PCR片段并经过测序后,得到至少有3-5个抗病候选基因存在于CT220标记处。随后,SPASSOVA等[13]在含有RFLP标识的CT220的长35250 bp的阳性BAC文库中筛选,分析发现2个属于CC-(NB-ARC)-LRR类型的抗性基因,而且同源性高达95%的候选抗性基因:Sw-5a和Sw-5b,经过转基因,证明Sw-5b能够使转基因烟草植株对TSWV的具备较强的抗性,但是Sw-5a则无抗性的产生。通过进一步的研究发现,依据氨基酸二级结构,Sw-5b划分为七个结构域,如下图。
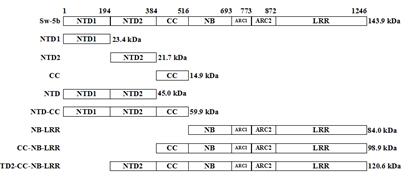
图1 Sw-5b各结构域划分示意图 番茄免疫受体蛋白Sw-5b与番茄斑萎病毒移动蛋白NSm互作分子机制研究(2):http://www.youerw.com/shengwu/lunwen_20856.html